Novel insights into the pathological development of dyslipidemia in patients with hypothyroidism
DOI:
https://doi.org/10.17305/bjbms.2021.6606Keywords:
Dyslipidemia, hypothyroidism, angiopoietin-like protein, fibroblast growth factor, PCSK9Abstract
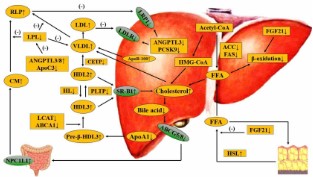
According to the previous reports, hypothyroidism has been shown to be strongly correlated with increased circulating concentrations of total cholesterol, low-density lipoprotein cholesterol (LDL-C), and triglycerides (TG). Notably, thyroid hormones are confirmed to modulate the production, clearance, and transformation process of cholesterol within circulation of mammals. Moreover, emerging evidence suggests that the thyroid-stimulating hormone could also participate in modulating serum lipid metabolism independently of thyroid hormones, which further induces the pathological development of dyslipidemia. However, the underlying mechanism is still not fully elucidated. Recently, several research studies have demonstrated that the pathogenic progression of hypothyroidism-related dyslipidemia might be correlated with the decreased serum concentrations of thyroid hormones and the increased serum concentrations of thyroid-stimulating hormones. Thus, this indicates that hypothyroidism could induce dyslipidemia and its related cardio-metabolic disorder diseases. In addition, several newly identified modulatory biomarkers, such as proprotein convertase subtilisin/kexin type 9 (PCSK9), angiopoietin-like protein (ANGPTLs), and fibroblast growth factors (FGFs), might play an important role in the regulation of dyslipidemia induced by hypothyroidism. Furthermore, under the status of hypothyroidism, significantly dysfunctional HDL particles could also be observed. In the current review, we summarized the recent knowledge of the relationship between the development of hypothyroidism with dyslipidemia. We also discussed the updated understanding of the mechanisms whereby hypothyroidism induces the risk and the development of dyslipidemia and cardio-metabolic diseases.
Citations
Downloads
Downloads
Additional Files
Published
License
Copyright (c) 2021 Xin Su, Xiang Chen, Hua Peng, Jingjin Song, Bin Wang, Xijie Wu

This work is licensed under a Creative Commons Attribution 4.0 International License.
How to Cite
Accepted 2021-11-02
Published 2022-06-01





