Inflammation-related cytokines and their roles in gastroenteropancreatic neuroendocrine neoplasms
DOI:
https://doi.org/10.17305/bjbms.2020.4471Keywords:
Inflammatory microenvironment, TNF-α, IL-6, IL-1β, IL-2, GEP-NENAbstract
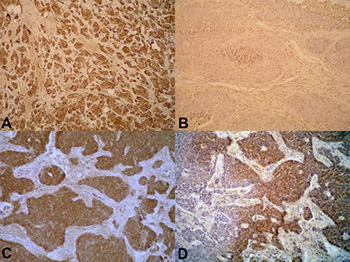
Proinflammatory counterworks are important at different stages of tumor development, particularly during invasion and metastasis. Immune cells and their signal molecules can influence all stages of tumor progression, as well as therapeutic intervention. Proinflammatory cytokines are known triggers of growth in gastroenteropancreatic neuroendocrine neoplasms (GEP-NENs). In this study, we explored the immunohistochemical expression of tumor necrosis factor alpha (TNF-α), interleukin 1 beta (IL-1β), IL-2, and IL-6 in tissues from 43 GEP-NEN patients with tumors of gastric, duodenal, ileal, appendiceal, and colonic origin. The immunohistochemical expression of TNF-α was increased in tumor groups with high proliferation rates (Ki-67; p = 0.034), as well as in those with higher tumor grades (p = 0.05). Moreover, the immunohistochemical expression of TNF-α positively correlated with death outcomes (p = 0.016). Expression of IL-6, IL-1β, and IL-2 displayed similar immunohistochemical expression patterns regardless of Ki-67, although the expression between the ILs differed. Most GEP-NENs had high levels of IL-6 and lower levels of IL-1β and IL-2. Although further comprehensive studies are required for a complete understanding of activated mechanisms in proinflammatory protumoral microenvironment of GEP-NENs, TNF-α is a potential marker in the prognosis of those tumors.
Citations
Downloads
Downloads
Additional Files
Published
How to Cite
Accepted 2020-01-21
Published 2020-11-02





