Effect of CCT137690 on long non-coding RNA expression profiles in MCF-7 and MDA-MB-231 cell lines
DOI:
https://doi.org/10.17305/bjbms.2019.4155Keywords:
Breast Neoplasms, CCT137690, MCF-7 Cells, MDA-MB-231, Long Non-coding RNAAbstract
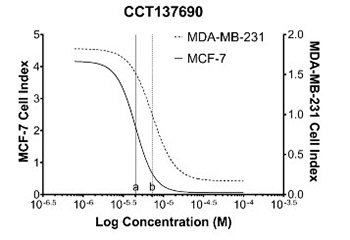
Long non-coding RNAs (lncRNAs) are involved in a range of biological processes, such as cellular differentiation, migration, apoptosis, invasion, proliferation, and transcriptional regulation. The aberrant expression of lncRNAs plays a significant role in several cancer types. Aurora kinases are increasingly expressed in various malignancies; accordingly, the inhibition of these enzymes may represent a novel approach for the treatment of various cancers. CCT137690, an Aurora kinase inhibitor, displays an anti-proliferative activity in human cancer cell lines. The aim of the present study was to investigate the anti-proliferative and cytotoxic effects of CCT137690 on estrogen receptor (ER)-positive human breast cancer cell line (MCF-7) and ER-negative human breast cancer cell line (MDA-MB-231). In addition, this study was targeted toward determining the changes induced in lncRNA expression levels following the initiation of Aurora kinase inhibitor treatment. The cytotoxic effects of CCT137690 were determined by means of the xCELLigence system. Furthermore, the anti-proliferative role of CCT137690 in breast cancer was investigated by checking the changes in lncRNA expression profiles using quantitative reverse-transcription polymerase chain reaction (qRT-PCR). The half-maximal inhibitory concentrations (IC50) of CCT137690 were determined as 4.5 μM (MCF-7) and 7.27 μM (MDA-MB-231). Several oncogenic lncRNAs (e.g., PRINS, HOXA1AS, and NCRMS) were downregulated in both ER-negative and ER-positive cell lines. On the other hand, tumor suppressor lncRNAs (e.g., DGCR5 and IGF2AS) were upregulated in the ER-positive cell line. After CCT137690 treatment, HOXA11AS and PCAT-14 lncRNAs were downregulated in the ER-positive cell lines. In addition, MER11C, SCA8, BC200, HOTAIR, PCAT-1, UCA1, SOX2OT, and HULC lncRNAs were downregulated in the ER-negative cell lines. The results of the present study indicated that Aurora kinase inhibitor CCT137690 could be a potential anti-cancer agent for breast cancer treatment.
Citations
Downloads
Downloads
Additional Files
Published
How to Cite
Accepted 2019-04-02
Published 2020-02-05





